白血病細胞が悪性度を維持しながら増殖するメカニズムを解明
2023年3月10日
公益財団法人 庄内地域産業振興センター
東京大学大学院新領域創成科学研究科
国立研究開発法人国立がん研究センター
研究成果のポイント
- 現行の治療法で治癒をもたらすことが難しい悪性度が高い白血病を引き起こすMOZ/MORF融合タンパク質が働くメカニズムを解明しました。
- 細胞分裂の前後でMOZ/MORF融合タンパク質が、CG配列を多く含む遺伝子プロモーターを活性化することで、悪性度を保ちながら無限増殖を引き起こすことを明らかにしました。
- MOZ/MORF融合タンパク質がCG配列含むDNAと結合する仕組みをタンパク質の立体構造レベルで明らかにしました。
- 本研究の成果によりMOZ/MORFを標的とした分子標的薬の開発が進み、白血病治療が発展することが期待されます。
概要
公益財団法人庄内地域産業振興センター(理事長:皆川治、鶴岡市末広町)/国立がん研究センター・鶴岡連携研究拠点がんメタボロミクス研究室(横山明彦チームリーダー)、東京大学大学院新領域創成科学研究科(金井昭教特任准教授)、米国のコロラド大学(Tatiana Kutateladze教授)及びカナダのラヴァル大学(Jacques Côté教授)の研究グループは、悪性度が高い白血病を引き起こすMOZ融合タンパク質*1やMORF融合タンパク質が働くメカニズムの一端を解明しました。MOZ/MORF融合タンパク質は細胞が分裂する時に、「自己複製*2」という特別な分裂様式を取ることで、細胞が悪性度を失わずにいつまでも増殖し続けるようになります。細胞は自己複製をすると、細胞分裂の前と後で同じ遺伝子セットを発現します。つまり細胞が自分と同じクローンを作る時に使う分裂様式が自己複製となります。正常な体においては自己複製する事ができるのは幹細胞と呼ばれる特別な細胞だけですが、異常なMOZ/MORF融合遺伝子が作られると、本来であれば自己複製しない造血細胞までもが自己複製するようになり、その結果がん細胞のクローンが無制限に生み出され、白血病を引き起こします。しかし、これまでMOZ/MORF融合タンパク質がどのようなメカニズムで自己複製を促進するのかわかっていませんでした。
MOZとMORFは非常によく似た構造を持つファミリータンパク質であり、ほぼ同じ働きをすると考えられています。MOZ/MORFにはウィングドヘリックスと呼ばれる構造が二つずつ含まれているのですが、その機能は不明でした。今回、研究グループはこの二つのウィングドヘリックス構造がDNAと結合することを発見しました。様々な配列のDNAと混ぜ合わせ、選択的に結合してくるDNAの配列を解析することで、一つ目のウィングドヘリックス(WH1)がメチル化されていないCG配列に特異的に結合することがわかりました。人のゲノムにあるほとんどの遺伝子は、プロモーター*3領域にCGに富む配列を持っています。プロモーター上のCG配列がメチル化されていないと、その遺伝子は発現する事ができますが、メチル化されると発現しなくなります。従って親細胞が発現していた遺伝子のプロモーターには多くの「メチル化されていないCG配列」が含まれています。MOZ/MORF融合タンパク質はWH1を介してメチル化されていないCG配列と結合することで、親細胞の時に発現していた遺伝子のプロモーターを認識し、転写を活性化します。これによって、細胞分裂前に発現していた遺伝子のプロモーターを細胞分裂後に再び活性化し、自己複製を促進すると考えられます。MOZ-TIF2融合タンパク質はWH1ドメインを失うと、プロモーターに結合できなくなり、白血病を引き起こさなくなりました。本研究はMOZ融合タンパク質がWH1を介してメチル化されていないCG配列と結合する事で、プロモーターに作用し、自己複製を促進することを明らかにしました。この結合は白血病化において必須のイベントであることから、創薬標的となる可能性があります。
本研究は、米国のコロラド大学及びカナダのラヴァル大学、そして日本の国立がん研究センター(理事長:中釜斉、東京都中央区)及び東京大学大学院新領域創成科学研究科(研究科長:出口敦、千葉県柏市)による国際共同研究であり、2023年2月8日に国際学術誌「Nature Communications」に掲載されました。
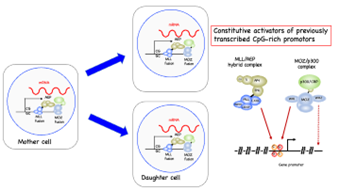
図1 MOZ融合タンパク質(MOZ fusion)やMLL融合タンパク質(MLL fusion)が自己複製を促進するメカニズム(左)とその分子メカニズムのモデル(右)。MOZ融合タンパク質は一つ目のウィングドヘリックス構造(WH1)を介して「メチル化されていないCG配列」に結合する。
背景
白血病は若年層で最も多く見られるがんであり、現行の治療法で治癒をもたらすことが難しい予後不良のタイプがあります。遺伝子異常の一種である「染色体転座」によって産みだされるMOZやMORF融合遺伝子は非常に強い発がんドライバー*4として機能し、予後不良の白血病を引き起こします。同じように予後不良の白血病を引き起こす遺伝子変異としてMLL融合遺伝子があります。我々はこれまでに、MLL融合タンパク質とMOZ融合タンパク質が同じゲノム上のプロモーターに結合する事を見出し(図1)、報告してきましたが(Miyamoto et al. 2020 Cell Reports 32:13;108200)、その分子基盤は不明でした。MLL融合タンパク質にはCXXCドメインというメチル化されていないCG配列に直接結合する機能ドメインがあり、それによってCG配列を含む様々な遺伝子プロモーターと結合することが明らかにされていました(図1、右)。一方で、MOZ/MORF融合タンパク質にはCXXCドメインはないため、どのようなメカニズムで遺伝子プロモーターを認識しているかは不明でした。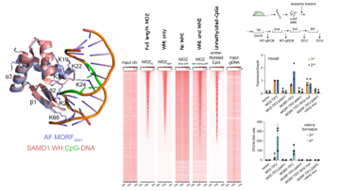 図2 MOZ/MORFのウィングドヘリックス構造の機能。左、MORFのWH1がメチル化されていない」CG配列と結合する構造。中央、ChIP-seq解析によって明らかになったMOZタンパク質のゲノム局在パターン。右、様々なMOZ/MORF融合タンパク質が正常な造血細胞を不死化する活性。MOZ/MORF融合タンパク質は標的遺伝子であるHOXA9を活性化して細胞を不死化する。
図2 MOZ/MORFのウィングドヘリックス構造の機能。左、MORFのWH1がメチル化されていない」CG配列と結合する構造。中央、ChIP-seq解析によって明らかになったMOZタンパク質のゲノム局在パターン。右、様々なMOZ/MORF融合タンパク質が正常な造血細胞を不死化する活性。MOZ/MORF融合タンパク質は標的遺伝子であるHOXA9を活性化して細胞を不死化する。
研究成果
米国のコロラド大学のKutateladze、カナダのラヴァル大学のCôté、国立がん研究センターの横山、東京大学の金井らの研究グループはMOZ/MORFタンパク質に含まれるウィングドヘリックスという構造に着目し、その機能を調べました。まず、研究グループはMORFに含まれる二つのウィングドヘリックス構造が両方ともDNAと結合する性質を持っており、特に一つ目のウィングドヘリックス(WH1)が遺伝子プロモーターとの結合に重要である事を見出しました。ウィングドヘリックス構造がDNAと結合する性質についてさらに詳しく解析していくと、WH1は「メチル化されていないCG配列」と結合し(図2左)、二つ目のウィングドヘリックスであるWH2は逆に「ATに富む配列」と結合する事がわかりました。これらのウィングドヘリックス構造がゲノム上の局在に与える影響をChIP-seq法*5と呼ばれる方法で解析したところ、WH1を持つタンパク質が「メチル化されていないCG配列」を多く含むゲノム領域に特異的に結合する事を示しました(図2、中央)。また、MOZ-TIF2融合遺伝子のWH1を欠損させた変異体はマウスの造血細胞に無限増殖能を与える事ができなくなっていたことから、WH1とCG配列の結合が白血病発症において必須のイベントであることが明らかになりました(図2右、MOZ-TIF2とMOZ-TIF2DWH1参照))。一方で、WH2を欠損させると、CG配列への結合能は保持されていましたが、遺伝子プロモーターへの結合の程度は減少しており、WH2もまたMOZ融合タンパク質がゲノムと結合する上で、補助的な役割を果たしている事が示唆されました(図1右)。実際、WH2の機能を欠損させたMORF融合タンパク質変異体は発がんドライバーとして十分な機能を発揮する事ができず、WH2を持つMORF融合タンパク質よりもやや弱くHOXA9遺伝子の発現を活性化し、培養を続けても最終的には造血細胞を不死化することはできませんでした(図2右、MORF-TIF2とMORF-TIF2 WH2 mut参照)。これらの結果はMOZ/MORF融合タンパク質が発がんドライバーとして機能するためには二つのウィングドヘリックスを介してDNAと強く結合する事が必要であり、この機能を阻害するような薬剤は分子標的薬となりうる事を示唆しました。
展望
本研究結果はMOZ/MORF融合タンパク質が造血細胞の異常な自己複製を引き起こす上で、ウィングドヘリックス構造を介したDNAとの結合が鍵となることを示しました。特に、WH1がメチル化されていないCG配列と結合するという知見はMOZ/MORF融合タンパク質が広範な遺伝子プロモーターを活性化し、親細胞の時に発現していたプロモーターを娘細胞においても活性化する分子基盤を説明するものでした。これらの知見はMOZ/MORFタンパク質が造血細胞の自己複製を制御する重要なタンパク質あることを示すとともに、創薬開発においても重要な分子標的である事を示しました。今後はMOZ/MORFを標的とした分子標的薬の開発が進み、白血病治療が発展することが期待されます。
発表論文
雑誌名
Nature Communications
タイトル
MORF and MOZ acetyltransferases target unmethylated CpG islands through the winged helix domain
著者
#Becht DC, #Klein BJ, #Kanai A, #Jang SM, Cox KL, Zhou BR, Phanor SK, Zhang Y1, Chen RW, Ebmeier CC, Lachance C, Galloy M, Fradet-Turcotte A, Bulyk ML, Bai Y, Poirier, MG, *Côté J, *Yokoyama A, and *Kutateladze TG.
#co-first author, *co-corresponding author
DOI
10.1038/s41467-023-36368-5
URL
https://www.nature.com/articles/s41467-023-36368-5(外部サイトにリンクします)
掲載日
2023年2月8日
用語解説
*1融合遺伝子/融合タンパク質
染色体は放射線などの影響で分断されると細胞内の修復メカニズムによって再結合するが、間違って元の染色体と異なる染色体断片同士が結合することで、遺伝子が再配列された染色体が生み出される現象を染色体転座という。その結果、二つの異なる遺伝子が融合した融合遺伝子が形成される。この遺伝子から発現されるタンパク質を融合タンパク質と呼ぶ。
*2自己複製
細胞が細胞分裂により1つ,もしくは2つの母細胞と同じ能力をもった娘細胞を生み出すこと
*3プロモーター
RNAをコードする遺伝子の先端部分のゲノム領域であり、その遺伝子の転写の起点となる。
*4 発がんドライバー
遺伝子の変異の内でがんの発症に関与する変異。
*5 ChIP-seq法
クロマチン免疫沈降法と呼ばれる手法で特定のタンパク質が結合するゲノムDNA断片を回収し他のち、DNAを次世代シーケンサーによって解析する事で、あるタンパク質がゲノム上に結合する領域を網羅的に同定する手法。
研究費
- がんメタボローム研究推進支援事業費補助金(山形県、鶴岡市)、横山明彦(代表)
- ENL変異型小児腎腫瘍の分子メカニズムの解明及び分子標的療法の開発 国際共同研究加速基金(国際共同研究強化(B))、横山明彦(代表)、金井昭教(分担)、R4-8年度、日本学術振興会
- AF10転座白血病における分子病態の解明及び新規治療法の開発、科学研究費助成事業 基盤研究(B)、横山明彦(代表)、金井昭教(分担)、R4-6年度、日本学術振興会
国立がん研究センター・鶴岡連携研究拠点がんメタボロミクス研究室について
国立がん研究センターのがんのメタボローム研究分野の研究拠点として、 山形県鶴岡市に2017年4月に設置。鶴岡連携研究拠点では、学校法人慶応義塾、慶應義塾大学先端生命科学研究所と連携して、メタボローム解析を活用した、がんの診断薬などの開発等に向けた研究を実施している。また企業との共同研究をより積極的に推進することにより、がんの分子基盤に基づいた新しい診断・治療法開発を進めている。
研究に関するお問い合せ先
国立研究開発法人国立がん研究センター・鶴岡連携研究拠点
がんメタボロミクス研究室 チームリーダー 横山明彦
〒997-0052 山形県鶴岡市覚岸寺字水上246番地2
鶴岡市先端研究産業支援センター内 がんメタボロミクス研究室
電話番号:0235-64-0980(事務室)、0235-64-1337(研究室)
FAX:0235-64-0981 Eメール:ayokoyam●ncc-tmc.jp
広報窓口
東京大学大学院新領域創成科学研究科
広報室
電話番号:04-7136-5450
Eメール:press●k.u-tokyo.ac.jp
国立研究開発法人国立がん研究センター
企画戦略局 広報企画室(柏キャンパス)
電話番号:04-7133-1111(代表) FAX:04-7130-0195
Eメール:ncc-admin●ncc.go.jp
関連ファイル
PDFファイルをご覧いただくには、Adobe Readerが必要です。Adobe Readerをお持ちでない方は、バナーのリンク先から無料ダウンロードしてください。