細胞内の「新型温度センサー」を発見!天然変性タンパク質が核内ストレス体の「るつぼ機能」を制御
大阪大学
北海道大学
国立研究開発法人国立がん研究センター
科学技術振興機構
発表のポイント
- ヒト細胞において熱ストレス応答を担う非膜オルガネラ「核内ストレス体」の温度感知機構を解明
- 特定の立体構造をもたない天然変性タンパク質PPP1R2が、温度変化を可逆的に感知する新型の「温度センサー」として機能することを新たに発見
- この働きにより、リン酸化酵素CLK1が温度依存的に核内ストレス体へ局在し、核内ストレス体が「るつぼ」として機能する仕組みを解明
- 細胞が環境変化に適応する仕組みやストレス関連疾患の研究発展に期待
概要
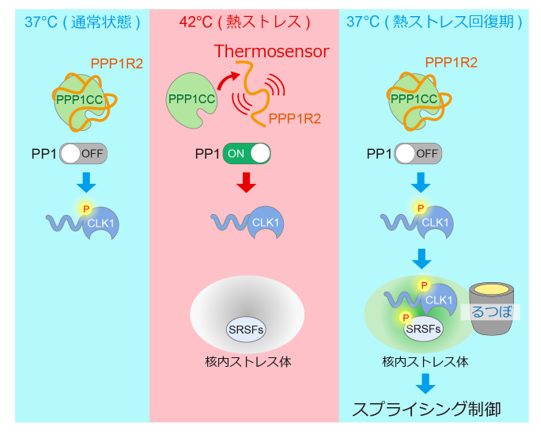
図1
天然変性タンパク質PPP1R2は温度センサーとして働き、CLK1のリン酸化と核内ストレス体 (nSB) への局在化を制御します。その結果、nSBによるスプライシング制御が温度依存的に調節されます。
大阪大学大学院生命機能研究科の上野剛志さん(研究当時:同理学研究科博士課程・日本学術振興会特別研究員 (DC2) 現:日本学術振興会特別研究員(PD))、二宮賢介特任講師、谷口一郎特任助教(常勤)、廣瀬哲郎教授(同理学研究科、先導的学際研究機構兼任)の研究グループは、国立がん研究センター研究所の足達俊吾部門長、北海道大学遺伝子病制御研究所の野田展生教授との共同研究によって、細胞内で温度を感知する新たな「温度センサー」を発見し、その情報が遺伝子発現制御へと変換される一連の仕組みを明らかにしました。
ヒト細胞は熱ストレスを受けると、核内に核内ストレス体(nSB)と呼ばれる膜のないオルガネラ(非膜オルガネラ)を形成します。nSBは熱ストレスから回復する過程で標的遺伝子の発現をRNAスプライシングの段階で制御することが知られていました。この過程では、熱ストレス時に脱リン酸化された不活性型スプライシング制御因子SRSFがnSB内に集積し、温度が正常に戻ったストレス回復期になると、SRSFのリン酸化酵素CLK1が取り込まれて、SRSFをnSB内で効率よくリン酸化し、再活性化します。このように、nSBはSRSFのリン酸化の酵素と基質を濃縮することで反応を促進する「るつぼ」として機能することが知られていました。しかし温度変化がどのようにCLK1のnSBへの集積を制御し、この「るつぼ機能」を作動させるのかは明らかになっていませんでした。
今回、研究グループは、CLK1が熱ストレス回復期にのみnSBへ集積する仕組みを解析し、CLK1のSer341リン酸化がこの局在制御の鍵であることを見出しました。さらに、このリン酸化状態がPP1(Protein Phosphatase 1)による脱リン酸化とRIOK2 (RIO Kinase 2) によるリン酸化によって制御されることを明らかにしました。
さらに、全体が特定の決まった立体構造をとらない天然変性タンパク質で、PP1の阻害因子であるPPP1R2が温度を感知する新たな「温度センサー」であることを発見しました。PPP1R2は高温条件でPP1から解離し、温度低下に伴って再結合することが判明しました。この可逆的な結合変化によってPP1活性が温度依存的に変化し、CLK1リン酸化状態の制御を介してnSBの「るつぼ」機能の切り替えを実現していることが示されました。
本研究は、天然変性タンパク質が温度変化を直接感知するセンサーとして機能することを初めて明らかにした成果であり、非膜オルガネラの機能制御への理解を深めるとともに、細胞の環境応答機構の理解に新たな視点をもたらします。本研究成果は、米国科学誌「Molecular Cell」に、7月17日(金曜日)午前1時(日本時間)に公開されました。
論文情報
雑誌名
Molecular Cell
タイトル
Thermo-Sensing Mechanisms of Splicing Control by Nuclear Stress Bodies
著者名
Tsuyoshi Ueno, Shungo Adachi, Ichiro Taniguchi, Nobuo N. Noda, Kensuke Ninomiya, and Tetsuro Hirose
DOI
https://doi.org/10.1016/j.molcel.2026.06.034(外部サイトにリンクします)
詳細
詳細は大阪大学のホームぺ―ジ(外部サイトにリンクします)をご覧ください。
お問い合わせ先
広報窓口
国立研究開発法人国立がん研究センター
企画戦略局 広報企画室
TEL:03-3542-2511(代表)
E-mail:ncc-admin●ncc.go.jp
関連ファイル
PDFファイルをご覧いただくには、Adobe Readerが必要です。Adobe Readerをお持ちでない方は、バナーのリンク先から無料ダウンロードしてください。